全國服務咨詢熱線:
自噬的研究背景
關于自噬的研究曾獲得過兩次諾貝爾生理學或醫(yī)學獎。是比利時科學家克里斯汀·德·迪夫(Christian de Duve)因發(fā)現(xiàn)了在細胞自噬機制中起到重要作用的溶酶體,獲得1974年諾貝爾獎;其次是日本大隅良典團隊因探明了細胞自噬的啟動機制,獲得2016年諾貝爾獎。2023年,國家自然科學基金委員會揭曉的今年國自然評審結(jié)果,自噬相關研究的中標數(shù)量超500項(來源:ZCOOL國自然項目查詢系統(tǒng))!
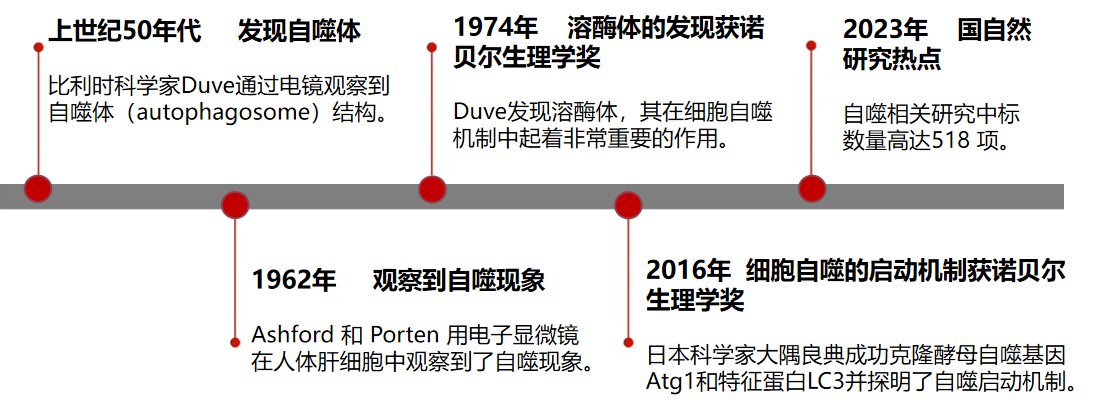
圖1 自噬研究進程
什么是自噬?
自噬是細胞在相關基因(autophagy related gene,Atg)的調(diào)控下利用溶酶體降解自身的細胞器和大分子物質(zhì)的過程,以此實現(xiàn)細胞本身的代謝需要和某些細胞器的更新,是真核細胞的生命現(xiàn)象。簡單來說就是細胞自己吃自己、廢物再利用。自噬現(xiàn)象在機體的生理和病理過程中都會發(fā)生,常見的病理誘導因素包括:營養(yǎng)缺乏、能量代謝異常、缺血、缺氧、病原體感染等。
細胞自噬主要有三種形式:微自噬(microautophagy)、巨自噬(macroautophagy,通常也稱作自噬)和分子伴侶介導的自噬(Chaperone-mediated autophagy,CMA),它們的主要不同之處在于產(chǎn)物(貨物)進入溶酶體的分子途徑(圖2)。
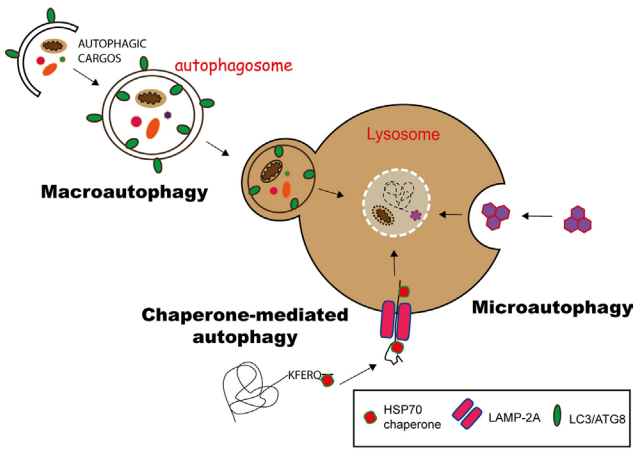
圖2 三種自噬途徑[1]
在分子伴侶介導的自噬(CMA)中,具有KFERQ樣基序的蛋白質(zhì)在Hsp70伴侶的幫助下通過LAMP-2A轉(zhuǎn)運蛋白轉(zhuǎn)運到溶酶體中。微自噬(microautophagy)涉及通過內(nèi)陷溶酶體膜來隔離底物,而在巨自噬(macroautophagy)中,底物被吞噬在稱為自噬小體的雙層膜小泡中,自噬小體隨后與溶酶小體融合,提供其內(nèi)容物以供降解。
自噬相關信號通路及關鍵蛋白
自噬過程受到不同自噬相關基因(autophagy-related gene,ATG)的調(diào)控, 據(jù)報道目前大約已有40個在酵母和哺乳動物中高度保守的自噬相關基因被發(fā)現(xiàn)。自噬發(fā)生需要經(jīng)過以下幾個階段:自噬前體形成、自噬前體延長包裹自噬的底物形成自噬泡(Phagophore)、細長自噬泡關閉形成自噬體(Autophagosome)、自噬體與溶酶體融合形成自噬溶酶體(Autolysosome)完成底物降解。
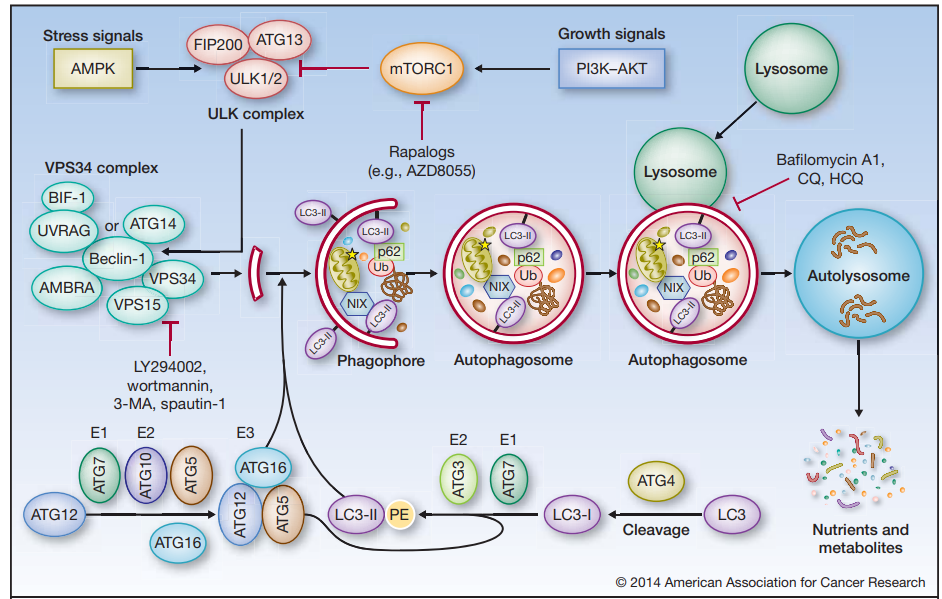
圖3 自噬過程[2]
典型的自噬(巨自噬)如圖3所示,在營養(yǎng)缺乏等應激情況下,AMPK的激活或mTOR的抑制導致ULK的激活,ULK使Beclin-1磷酸化,導致VPS34的激活和吞噬細胞的形成。ULK在與FIP200和ATG13的復合體中發(fā)揮作用,而VPS34的功能需要一個調(diào)節(jié)亞基VPS15(P150)和Beclin-1,后者進一步調(diào)節(jié)其他調(diào)節(jié)因子如AMBRA、ATG14、UVRAG和BIF-1的聯(lián)系。多種ATG蛋白,如ATG5和ATG7,構(gòu)成了兩個“泛素樣結(jié)合系統(tǒng)",它們催化形成磷脂酰乙醇胺(PE)連接的LC3(LC3-II),并將其正確地結(jié)合到吞噬體膜上。細長自噬泡的關閉標志著成熟自噬體的形成,自噬體最終與溶酶體融合,導致貨物降解、營養(yǎng)物質(zhì)和代謝物的循環(huán)。
表1自噬不同階段關鍵蛋白及作用
階段 | 蛋白名稱 | 功能 | |
起始及吞噬泡成核 | ULK1 復合體 | ATG1/ULK1 | 通過自噬機制的磷酸化組分啟動自噬; |
ATG13 | 介導ULK1和FIP200之間相互作用的接頭; | ||
ATG17/FIP200 | ULK1復合物的組成部分; | ||
PI3KC3 復合體 | ATG6/Beclin1 | 促進PI3KC3-C1復合體形成; | |
VPS34 | 促進PI3KC3-C1復合體的形成,ULK1的磷酸化位點,穩(wěn)定ULK1復合物。 | ||
ATG14 | ULK1的磷酸化位點; | ||
VPS15 | 絲氨酸/蘇氨酸激酶; | ||
AMBRA | Beclin1的相互作用蛋白 | ||
吞噬泡延伸 | ATG8/LC3-PE | ATG8/LC3 | 以LC3-I和LC3-Ⅱ兩種形式存在;參與自噬體膜的形成,與自噬體膜表面的PE結(jié)合,可作為自噬體的標記分子 |
ATG7 | E1樣酶; | ||
ATG12-ATG5-ATG16復合體 | ATG12&ATG5&ATG16 | ATG5直接結(jié)合膜,這種膜結(jié)合受ATG12的負調(diào)節(jié),但被ATG16激活。 | |
貨物包裹 | 關鍵因子 | Ubiquitin | 標記貨物 |
SQSTM1/p62 | 同OPTN、NBR1和NDP52,都是自噬受體,識別貨物。 | ||
PE-LC3 | 與自噬受體的相互作用; | ||
Rabs | Rab1&Rab11 | 參與自噬體的生物合成 | |
Rab7 | 促進啟動階段自噬體與溶酶體的融合; | ||
自噬體與溶酶體融合 | HOPS 復合體 | VPS11 | 與VPS16、VPS18、VPS33、VPS39和VPS41共同識別SNAREs復合體并調(diào)節(jié)其組裝, |
SNAREs 復合體 | STX17 | 介導自噬體與溶酶體融合,也參與早期自噬體的成熟。 | |
SNAP29 | 介導自噬體與溶酶體融合。 | ||
YKT6 | 與STX17-SNAP29形成復合體,介導自噬體與溶酶體的融合。 | ||
VAMP7/8 | 位于溶酶體,介導自噬體與溶酶體的融合。 | ||
細胞自噬研究策略
1、電鏡觀察自噬體的形成——自噬檢測金標準r
自噬體屬于亞細胞結(jié)構(gòu),需要使用透射電子顯微鏡觀察自噬體的形態(tài)。電鏡檢測自噬主要是基于辨認自噬體結(jié)構(gòu)特征(表2)。
表2 自噬不同階段形態(tài)特征
自噬階段 | 特征 |
自噬泡(Phagophore) | 新月狀或杯狀,雙層或多層膜,有包繞胞漿成分的趨勢。 |
自噬體(Autophagosome) | 雙層或多層膜的液泡狀結(jié)構(gòu),內(nèi)含胞漿成分,如線粒體、內(nèi)質(zhì)網(wǎng)、核糖體等。 |
自噬溶酶體(Autolysosome) | 單層膜,胞漿成分已降解。 |
依照上述描述,能夠確認大部分自噬性囊泡,但有以下問題需要注意:
1)是否存在雙層或多層膜不能作為識別自噬體的依據(jù)(普通透射電鏡切片在制備過程中的處理可影響膜脂質(zhì)成分,切片上的自噬體并非都是兩層膜結(jié)構(gòu),可能僅有一層膜,也可能是多層膜,有時膜結(jié)構(gòu)可能因為脂質(zhì)被提取而無法辨認);
2)線粒體也含雙層膜,腫脹或含有沉淀物的線粒體外觀與自噬體形態(tài)相似,但仔細觀察,線粒體雙層膜間距一般較小且均一,線粒體內(nèi)膜折疊形成嵴,而自噬體內(nèi)膜不會折疊;
3)粗面內(nèi)質(zhì)網(wǎng)有時會包繞線粒體等細胞器,容易誤認為自噬體,根據(jù)內(nèi)質(zhì)網(wǎng)上有核糖體而自噬體膜上無核糖體可鑒別;
4)切片上的電子密度低或空泡有時也會被誤認為自噬囊泡。
2、Western blot檢測自噬相關蛋白
在進行自噬研究時,提取蛋白進行Western Blot,檢測LC3-I和LC3-II幾乎是必做的實驗。細胞內(nèi)存在兩種形式的LC3蛋白:LC3-Ⅰ和LC3-Ⅱ。LC3蛋白在合成后其C端即被Atg4蛋白酶切割變成LC3-Ⅰ,分布于細胞漿內(nèi)。當自噬體形成后,LC3-Ⅰ和磷脂酰乙醇胺(phosphatidylethanolamine, PE)偶聯(lián)形成LC3-Ⅱ并定位于自噬體內(nèi)膜和外膜,并穩(wěn)定地保留在自噬體膜上直到與溶酶體融合。因此,LC3-Ⅱ被用來作為自噬體的標記,其表達水平在某種程度上反映了自噬體的數(shù)量[3]。LC3-I/II的形成和降解是一個動態(tài)過程,瞬時LC3-Ⅱ表達不能反映自噬程度,需配合使用工具化合物分析自噬變化。
除了檢測自噬標志物LC3的轉(zhuǎn)換(LC3-II/LC3-I)外,還可以用Western Blot實驗檢測Beclin1、P62等自噬相關蛋白的表達情況。Western blot操作包括:總蛋白提取→蛋白含量測定→SDS-PAGE電泳→電轉(zhuǎn)→封閉→抗體孵育→化學發(fā)光成像。就操作難度而言,Western blot在一眾實驗中排名靠前,WB實驗具體步驟可參考我們往期分享。
3、自噬的實驗性調(diào)控——工具化合物
通過人為的干預來激活或者抑制自噬功能后觀察細胞行為或效應分子的變化能夠使研究結(jié)論更具有說服力。目前,常用的抑制自噬及誘導自噬的藥物及機制如表3所示。需要注意的是,這些工具藥普遍存在的缺陷就是特異性不強,在抑制自噬的同時對細胞其他代謝過程也可能會有影響。如3-MA抑制ClassⅢ PI3K的同時對ClassⅠ PI3K同樣也有抑制作用,繼而抑制Akt/mTOR通路,激活自噬。
表3 細胞自噬常用的激活劑和抑制劑
類別 | 工具化合物 | 作用原理 |
自噬 激活劑 | Metformin hydrochloride | AMPK激活劑 |
Rapamycin、Torin-1、Torkinib、AZD-8055 | mTOR 抑制劑 | |
ABT-737 | 競爭性破壞Beclin-1/ Bcl-2/Bcl-XL間的相互作用 | |
自噬 抑制劑 | 3-MA、LY 294002、Wortmannin、Spautin-1 | PI3K抑制劑,抑制自噬泡的形成 |
Nocodazole、Bafilomycin A1 | 抑制自噬體與溶酶體的融合 | |
Aloxistatin、Pepstatin A | 抑制溶酶體蛋白酶活性 |
4、應用 GFP-LC3 轉(zhuǎn)基因小鼠實現(xiàn)體內(nèi)自噬的實時監(jiān)測
Tian等[4]利用GFP-LC3轉(zhuǎn)基因小鼠,借助于體內(nèi)成像技術(shù)經(jīng)顱骨檢測實驗性腦卒中后缺血腦組織區(qū)域GFP熒光強度,與體外自噬成像、自噬抑制劑3-甲基腺嘌呤(3-MA)檢測、蛋白印跡分析、免疫組織化學、免疫熒光等分析方法結(jié)果相似。該研究提供了活體自噬檢測的證據(jù),展示了一種在活體動物模型中檢測體內(nèi)自噬的方法,此方法對監(jiān)測活體中風患者體內(nèi)的自噬過程以及闡明自噬在腦缺血和其他神經(jīng)疾病中的詳細作用具有重要價值。
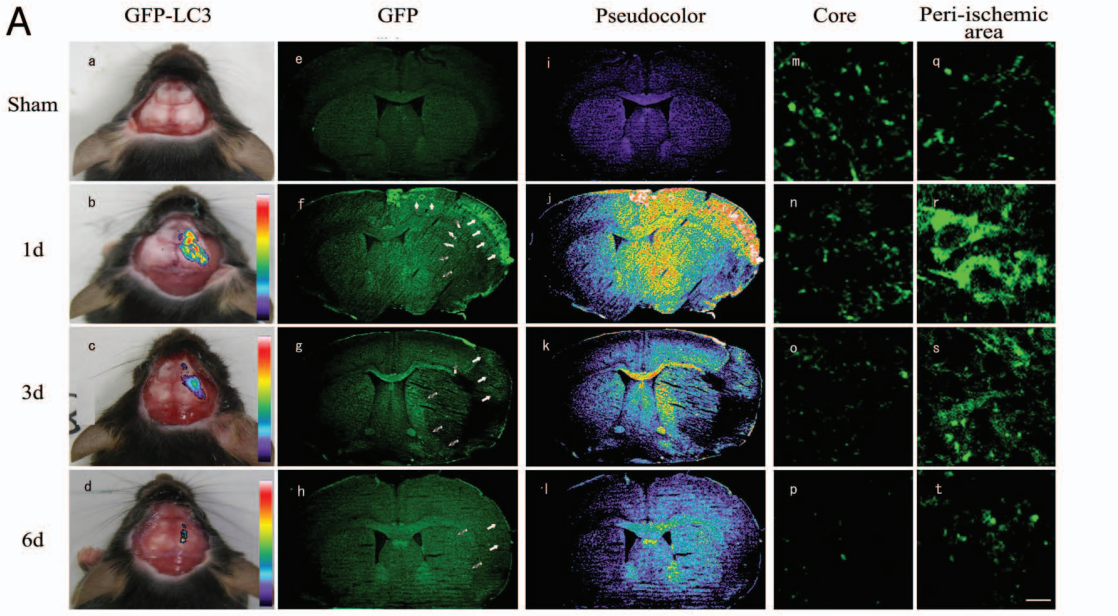
圖4 腦缺血區(qū)頭部活體成像監(jiān)測GFP熒光信號
自噬研究已成為當前生命科學研究的熱點,揭示自噬的發(fā)生機制、自噬與疾病發(fā)生的關系對預防與治療多種人類重大疾病具有重要意義。然而,至今并沒有非常準確可靠的自噬功能檢測和監(jiān)控方法,我們在進行自噬相關研究時應結(jié)合不同的檢測原理,借助不同的檢測技術(shù)進行分析,體內(nèi)外結(jié)果互相驗證以避免單一技術(shù)的局限性。
參考文獻
[1] Duraes F V , Jennifer N , Juan D ,et al.Macroautophagy in Endogenous Processing of Self- andPathogen-Derived Antigens for MHC Class II Presentation[J].Frontiers in Immunology, 2015, 6. DOI:10.3389/fimmu.2015.00459.
[2] Michelle Cicchini; Vassiliki Karantza; Bing Xia.Molecular Pathways: Autophagy in Cancer-A Matter of Timing and Context[J].Clinical Cancer Research.2015,21 (3): 498-
[3] Kimura S, Fujita N, Noda T, et al. Monitoring autophagy inmammalian cultured cells through the dynamics of LC3. Methods Enzymol, 2009, 452: 1-12.
[4] Tian F, Deguchi K, Yamashita T, et al. In vivo imaging of autophagy in a mouse stroke model. Autophagy, 2010, 6(8): 1107-1114.
自噬激活劑:
貨號 | 產(chǎn)品名稱 | CAS號 | 規(guī)格 |
abs810030 | Rapamycin | 53123-88-9 | 100 mg |
abs810772 | AZD-8055 | 1009298-09-2 | 50 mg |
abs812114 | ABT-737 | 852808-04-9 | 50 mg |
abs812923 | Torin-1 | 1222998-36-8 | 20 mg |
abs817883 | Metformin hydrochloride | 1115-70-4 | 100 mg |
abs810030 | Rapamycin | 53123-88-9 | 100 mg |
自噬抑制劑:
貨號 | 產(chǎn)品名稱 | CAS號 | 規(guī)格 |
abs810001 | LY 294002 | 154447-36-6 | 100 mg |
abs810575 | 3-MA | 5142-23-4 | 200 mg |
abs817875 | Wortmannin | 19545-26-7 | 10 mg |
abs817886 | Nocodazole | 31430-18-9 | 100 mg |
abs817902 | Pepstatin A | 26305-03-3 | 50 mg |
abs817918 | Aloxistatin | 88321-09-9 | 25 mg |
自噬相關抗體:
貨號 | 產(chǎn)品名稱 | 規(guī)格 |
abs103336 | Rabbit anti-Cleaved LC3A Polyclonal Antibody | 50uL |
abs113447 | Rabbit anti-Cleaved LC3B Polyclonal Antibody | 50uL |
abs124528 | Rabbit anti-Nucleoporin p62 Polyclonal Antibody | 50uL |
abs131205 | Rabbit anti-Beclin 1 Polyclonal Antibody | 50? |
abs158317 | Mouse anti-LC3A/B Monoclonal Antibody(3E9) | 100uL |
abs158462 | Mouse anti-LC3A Monoclonal Antibody(8F5) | 100uL |
溫馨提示:absin所有產(chǎn)品僅用于科學研究,請勿藥物、家用或其他用途。
好消息!Absin文獻獎勵重磅升級!

Absin特色產(chǎn)品線:
WB相關:ECL發(fā)光液、預染marker、預制膠;IHC相關:二抗試劑盒、組化筆;IP/CoIP試劑盒;激動劑/抑制劑;血清、BSA、蛋白酶K、CTB、TTX、CEE;凋亡試劑盒;呼吸爆發(fā)試劑盒;ELISA試劑盒;重組蛋白;抗體: 二抗、標簽抗體、對照抗體;定制服務(抗體/多肽/蛋白/標記/檢測)...

電話
微信掃一掃